The primary features of cancer are maintained via intrinsically modified metabolic activity, which is characterized by enhanced nutrient supply, energy production, and biosynthetic activity to synthesize a variety of macromolecular components during each passage through the cell cycle.
Glutamine is the most abundant amino acid (building block of protein) in the body. Its main storage site is in the musculature; where about 60% of all the unbound amino acids are glutamine (glutamine makes up a smaller percentage of muscle protein, the main bound form). Glutamine has been called a “conditionally essential” nutrient, because it is non-essential in normal situations (meaning that the body can normally synthesize what it needs; not required in the diet), but in severe illness or injury becomes insufficient (there is then a need for supplementation from the diet or other sources). Glutamine is manufactured in the body from glutamate and ammonia by the enzyme glutamine synthetase; the process takes place mainly in the skeletal muscles.
Glutamine is crucial for many aspects of healthy body function, including maintenance of optimal antioxidant status, building and maintenance of muscle tissue, maintenance of optimal immune function, and repair and maintenance of intestinal tissue. It is important for removing excess ammonia (a common waste product in the body). It also appears to be needed for normal brain function and digestion. When the body is stressed (from injuries, infections, burns, trauma, or surgical procedures), it releases the hormone cortisol into the bloodstream. High levels of cortisol can lower body’s stores of glutamine. However, because it has long been classified as “non-essential” little attention has been paid to its importance.
The role of glutamine in cancer has been a topic of recent interest. On the one hand, glutamine seems to be the ideal treatment for the cachexia that accompanies tumor growth and the adverse reactions to chemotherapy. Many people with cancer have low levels of glutamine. For this reason, some researchers speculate that glutamine may be helpful when added to conventional cancer treatment for some people. Supplemental glutamine is often given to malnourished cancer patients undergoing chemotherapy or radiation treatments and sometimes used in patients undergoing bone marrow transplants.
On the other hand, it has been found that tumor cells are capable of efficiently transporting glutamine and that this is one of their major respiratory fuels. The connection of glutamine to the musculature is of special interest. The amount of glutamine in reserve for release as needed is directly related to muscle mass: more muscle mass means more glutamine is available for metabolic processes. Therefore, the question has been raised as to whether administering glutamine might not also be beneficial to the cancer.
More tumor mass means more glutamine is available for metabolic processes. The rapidly dividing cells such as white blood cells, intestine cells and cancer cells have a requirement for glutamine as a fuel. As the cancer develops, it can compete with the rest of the body for the glutamine supply. The body’s metabolism is forced to shift into high absorption of dietary glutamine and production of glutamine by the liver and other tissues. The following studies suggest that glutamine may actually stimulate growth of tumors.
Glutamine addiction: a new therapeutic target in cancer.
Glucose and glutamine are absolutely required for cell division that feed into multiple pathways required to support cell growth. In actively growing cells, most of the glucose is secreted as lactate; the synthesis of lactate generates ATP required to maintain intracellular bioenergetics. The remaining glucose enters TCA cycle where it is metabolized to citrate. Within mitochondria of rapidly growing cells, most of the glucose-derived citrate is exported to support the synthesis of fatty acids required for cell growth. High citrate efflux from the mitochondria “empties” the TCA cycle. Glutamine “refills” the TCA cycle by a process termed anapleurosis
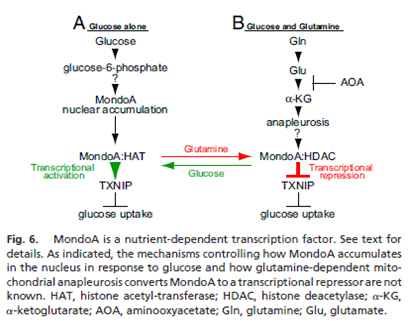
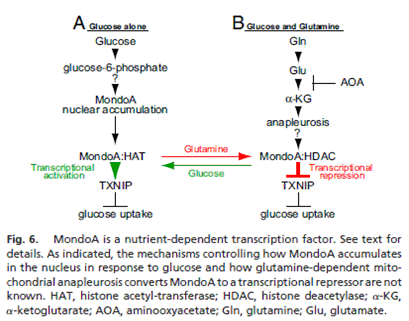
MondoA is a transcription factor. MondoA and its binding partner Mlx localize to the cytoplasm, suggesting tight regulation of their nuclear function. A key MondoA effector in this regulatory circuit is TXNIP (thioredoxin-interacting protein). Repression of TXNIP stimulates glucose uptake and aerobic glycolysis and also blocks fatty acid oxidation, which are features of rapidly dividing cells. Further, TXNIP reduction likely also drives cell division and survival more directly by triggering the degradation of the cyclin-dependent kinase inhibitor p27 and inhibition of the PTEN tumor suppressor. Glutamine triggers MondoA-dependent repression of TXNIP. Glutamine, via mitochondrial anapleurosis, converts MondoA:Mlx from a transcriptional activator to a transcriptional repressor. Rather than controlling MondoA:Mlx function indirectly by modulating subcellular localization or promoter occupancy, glutamine triggers the recruitment of an HDAC-dependent corepressor to promoter-bound MondoA:Mlx complexes.

The Mondo and Myc families may share some biological functions; it is likely each family is under distinct regulatory control. Cells overexpressing a conditional allele of the Myc are hypersensitive to glutamine deprivation and die when glutamine is removed from the medium. This glutamine “addiction” may result because Myc stimulates glutaminolysis by activating, directly or indirectly, the expression of genes involved in glutamine uptake and utilization
This metabolic shift in cancer cells, as compared with non-proliferating cells, involves aberrant activation of aerobic glycolysis, de novo lipid biosynthesis and glutamine-dependent anaplerosis to fuel robust cell growth and proliferation.
Blockade of glutamine metabolism also restricts glucose uptake. Inhibiting glutaminolysis might be efficacious in restricting cancer cell growth by limiting the availability of one key nutrient required for cell growth. As such, the combined inhibition of glycolysis and glutaminolysis may be especially effective at driving apoptosis of highly glycolytic tumor cells.

However, attempting to block tumor growth by starving it for glutamine is not practical, as the body will be starved of glutamine first. Using glutamine analogs that will be taken up by the tumors in place of glutamine but which fail to nourish the tumor have been developed, but have not been found to be viable treatments as yet. Instead, it has been suggested that cancer patients be given natural agents that inhibit TCA enzymes. Inhibition of TCA enzymes by SinnolZym is an attractive approach to sensitize cells to inhibition of glycolysis. SinnolZym activates AMPK, a key sensor of cellular energy status. AMPK activity is an important contributing factor in the reduced mitochondrial function and dysregulated intracellular lipid metabolism. LKB1 is a tumor suppressor protein. AMPK is an essential mediator of the tumor suppressor LKB1. AMPK can also be activated by LKB1.
AMPK as a metabolic tumor suppressor: control of metabolism and cell growth.
AMP-activated protein kinase in metabolic control and insulin signaling.
Dialogue between LKB1 and AMPK: a hot topic at the cellular pole.
LKB1 and AMPK family signaling: the intimate link between cell polarity and energy metabolism.
Important role of the LKB1-AMPK pathway in suppressing tumorigenesis in PTEN-deficient mice.
Thiazolidinediones (TZDs), synthetic insulin-sensitizing antidiabetic drugs that include troglitazone, pioglitazone, and rosiglitazone, increase P27. P27 inhibits growth and transformation of cancer cells. This response is independent of PPARgamma activation induced by TZDs. In alternative non toxic cancer treatment, SinnolZym and TZDs combination therapy has shown powerful synergistic antitumor effects against breast and gastric cancer. Such combination therapy may be most useful in the treatment of most cases cancer and leukemia.
Mechanisms by which thiazolidinediones induce anti-cancer effects in cancers in digestive organs.


{kind=link}
{kind=link}